CRISPR/Cas9技术在节肢动物中的应用研究
作者: 黄津伟 胡乐贞 齐佳辰 孙金生 李冉
摘要 CRSPR/Cas9系统是基于RNA的适应性免疫系统,广泛存在于细菌和古细菌中。其特点在于单个蛋白质Cas9与两个短RNA序列结合后,可作为位点特异性核酸内切酶在体内或体外发挥作用。与传统的由核酸酶介导的DNA编辑技术不同,CRISPR/Cas9对DNA的识别不是由蛋白质指定的,而是由 20-nt向导RNA(guide RNA)序列指定的。CRISPR/Cas9系统由于其设计过程和使用过程简单高效而被广泛应用。总结CRSPR/Cas9系统作用机理、导入方法及其在节肢动物中不同类群及不同基因的应用进展,以期为重要农业及经济节肢动物的功能基因研究与遗传育种研究提供有价值的参考。
关键词 CRISPR/Cas9;导入方法;节肢动物;基因编辑
中图分类号 Q78 文献标识码 A 文章编号 0517-6611(2022)01-0001-07
doi:10.3969/j.issn.0517-6611.2022.01.001
Research on the Application of CRISPR/Cas9 Technology in Arthropods
HUANG Jin-wei, HU Le-zhen, QI Jia-chen et al
(College of Life Sciences,Tianjin Normal University, Tianjin 300387)
Abstract The CRISPR/Cas9 system is an RNA-based adaptive immune system which exists in bacteria and archaea. Its characteristic is that following the compound of a single protein Cas9 with two short RNA, it can act as a site-specific endonuclease in vivo/in vitro. Unlike traditional nuclease-mediated DNA editing technology, the recognition of DNA by CRISPR-Cas9 is not specified by the protein, but by the 20-nt guide RNA (guide RNA) sequence. The CRISPR-Cas9 system is widely used due to its simplicity in design, use and high efficiency. Summarizes the mechanism of CRSPR/Cas9 system, the introduction method and the application progress of different groups and different genes in arthropods, in order to provide a valuable reference for the research of functional genes and genetic breeding of important agricultural and economic arthropods.
Key words CRISPR/Cas9;Introducing method;Arthropod;Gene editing
基金项目 “蓝色粮仓”国家重点研发计划子课题(2018YFD0901301)。
作者简介 黄津伟(1997—),女,天津人,硕士研究生,研究方向:水生生物学。
通信作者,孙金生,研究员,从事重要水产动物疾病的发生和免疫防治研究;李冉,副教授,从事重要水产动物生长生理及分子育种研究。
收稿日期 2021-08-01;修回日期 2021-08-23
基因组编辑技术是研究各种生物体靶基因功能的有效手段,在CRISPR/Cas9系统被发现之前,锌指核酸酶(zinc finger nucleases,ZFNs)和转录激活因子样效应核酸酶(transcription activator-like effector nucleases,TALENs)技术常用于基因编辑,这2种技术均可用于设计DNA结合域,可以有效地识别和修改基因序列,被广泛应用于各个领域。然而锌指核酸酶和转录激活因子样效应核酸酶需要使用多种核酸酶,核酸酶的脱靶效应会导致细胞毒性,且使用方法较为复杂[1],因此这2个基因组编辑系统最近被CRISPR/Cas9系统取代。CRISPR/Cas9系统进行基因编辑简单高效,已被广泛用于哺乳动物。
节肢动物物种繁多,包括与农林渔业相关的诸多物种,了解相关物种的基因功能从而在农林渔业中合理应用是非常重要且必要的。CRISPR/Cas9已应用于节肢动物的部分物种,笔者对已成功应用的物种及研究进展进行总结,以期为重要农业及经济节肢动物的功能基因研究与遗传育种研究提供有价值的参考。
1 CRISPR/Cas系统
1.1 CRISPR/Cas的作用机理及类型
CRISPR/Cas(Clustered Regularly Interspaced Short Palindromic Repeats/ CRISPR associate proteins)系统最早发现于细菌内,是以RNA为基础的适应性免疫系统,主要防御外来核酸的侵染,维持细菌自身正常生命代谢活动[2]。CRISPR/Cas系统由反式激活CRISPR RNA基因(tracrRNA)、Cas操纵子和CRISPR DNA区域3部分组成,其中CRISPR由一组重复间隔序列及多个非重复间隔序列组成,非重复间隔序列是早先侵入细菌内的病原体核酸片段经过不断进化形成的。tracrRNA作为一种独特的非编码RNA,与CRISPR间隔序列具有同源性。Cas操纵子目前已发现有多种,如Cas9、Cas1、Cas2和Csn2,可经转录翻译生成Cas蛋白(核酸酶)。当侵染过的病毒再次入侵细胞,CRISPR DNA区域转录生成前体CRISPR RNA(pre-crRNA),tracrRNA与pre-crRNA通过碱基互补配对及RNA酶的作用形成成熟的crRNA[3]。crRNA与tracrRNA组成单链导向RNA(sgRNA),并利用细菌中的核酸酶进一步修饰crRNA的5′端,使其长度最终减少到20 nt。当crRNA下游存在特定的前间区序列邻近基序(PAM)时,Cas蛋白将被引导至入侵病原体DNA与crRNA互补的位点处进行切割,完成免疫过程。基于上述过程,有研究者提出可以在体外合成sgRNA模拟体内crRNA和tracrRNA的结构,并与Cas9结合形成CRISPR/Cas9系统。Jinek等[4-5]研究出利用sgRNA对特定DNA进行剪切的核酸酶系统,首次证明了CRISPR/Cas9可以被应用于基因工程。Mali等[6-7]创建CRISPR/Cas9系统并应用于基因编辑。由于其设计使用较为简单且有效,CRISPR/Cas系统得到了生物学届和医学界的广泛关注,并广泛应用于基因编辑、转录扰动、表观遗传调节和基因组成像等相关研究[8]。根据目前CRISPR/Cas位点的分类[9],CRISPR系统被分为6种不同的类型(I~VI),各类型都使用特定的Cas蛋白和crRNA。相比其他系统来说,II型系统的结构最简单,即CRISPR/Cas9系统。
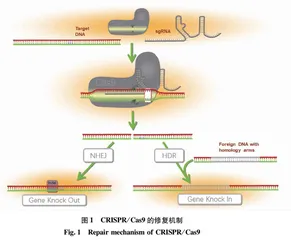
1.2 CRISPR/Cas9的修复机制
Cas9蛋白包含2个结构域,其中HNH结构域可剪切与sgRNA互补的靶DNA链(目标链),RuvC结构域则剪切另外一条核酸链,造成目标DNA的双链断裂(double-strand DNA break,DSB)。双链断裂后细胞内会启动修复机制,细胞在不同的分裂时期会启动不同的修复机制。每个细胞分裂时期都可以发生非同源修复机制(nonhomologous end joining,NHEJ),这种机制不需要同源模板存在。DNA双链断裂后修复酶会对断裂末端进行酶切修饰,便于DNA连接酶将断裂的两端DNA重新连接在一起,这种修复方式易在断裂位点出现碱基插入或缺失,且通常会产生移码突变,从而导致蛋白质翻译的提前终止,转录物降解或蛋白质丢失。另一种细胞修复机制只能发生在细胞分裂间期,它需要同源修复模板,称为同源修复机制(homology-directed repair,HDR),模板可以是ssDNA或dsDNA,在导入CRISPR/Cas9的同时,需要提供带同源臂的供体DNA作为整合模板,其可以携带外源DNA序列从而引入新的基因。CRISPR/Cas9系统也是当前应用最多的系统可以利用非同源修复机制使目的基因转录失败或使目的蛋白失去功能,进而实现基因敲除,在哺乳动物中的成功率较高,Cas9中的HNH结构域可以利用多个sgRNA通过非同源修复机制实现一个试验中多个靶基因(序列较短)的突变[7,10],也可通过引入2个sgRNA敲降较长的基因组片段。
1.3 CRISPR/Cas9系统的导入方法
1.3.1 CRISPR/Cas9系统的封装方式。
目前导入CRISPR/Cas9的方式有很多,分别可以mRNA、DNA、蛋白-核酸的形式引入。在RNA水平上,2015年Hruscha等[11]将sgRNA和Cas9 mRNA通过显微注射导入到斑马鱼胚胎中,对斑马鱼基因组进行编辑;在DNA水平上,主要通过质粒或病毒介导DNA导入;在蛋白质-核酸水平上,通常在体外表达Cas9蛋白,并与sgRNA孵育结合形成Cas9蛋白-sgRNA复合体后将其导入受精卵或细胞中进行基因编辑[12-13]。
1.3.2 显微注射法。
显微注射法是将外源基因导入胚胎的最常见且最直接的方法,在节肢动物的模式生物中应用较为广泛,也是当下CRISPR/Cas9导入的主要技术。但显微注射技术仍在应用方面存在一些阻碍:例如部分节肢动物的卵膜会在排卵后很快变硬导致在注射时胚胎破裂,由于卵细胞内外渗透压不同造成注射时内容物的外流等。目前研究人员针对不同情况已经提出了相应的解决方法,对于卵膜硬化问题,解决的方法是选择合适的时间,在产卵后应立即收集胚胎,将卵置于某些试剂(如蔗糖、次氯酸盐)中[13-14]或置于冰上,防止卵膜变硬;而对于卵细胞渗透压差异问题,平衡渗透压是最有效的解决方法,海洋动物的卵或胚胎可以放在海水中[15];探索合适的培养条件,如Hiruta等[16]提出水蚤中分离的胚胎培养于2%琼脂、60 mmol/L蔗糖的培养基上最有利于显微注射。虽然显微注射技术目前应用得相对广泛,但由于需要精密昂贵的注射设备、高难度的操作技术且成活率较低,研究者们在积极探求更优的技术方法。
1.3.3 精子介导法。
1971年首次报道了精子能携带异源基因整合到受精卵中[17],1989年Lavitrano等[18]将外源DNA与小鼠精子在等渗溶液中孵育15 min,在后代中检测出外源基因被带入受精卵,证明精子细胞具有吸收外源DNA并将其携带到卵细胞的能力。但是,由于精子是成熟的生殖细胞,无法在分裂之前选择成功整合外源DNA的细胞,即使精子进入卵细胞后在受精卵或子一代中成功转入外源基因,也可能是以染色体外形式存在,而不是整合到宿主基因组中[19]。2008年Wu等[20]将109个精子和50~300 mg DNA在体外17 ℃孵育,后经人工授精,检测到仔猪存在转基因个体,但70~90 d龄的仔猪均未检测到外源DNA,表明精子摄取DNA仅在早期传递给后代,研究人员推测是由于早期细胞分裂复制过程导致外援DNA丢失。为了优化精子介导法,研究者进行了多种尝试,例如使用线性化质粒[18]、通过转染试剂优化DNA转移[21],并且发现精子摄取DNA效率与精子的运动性高度相关[20]。目前精子介导基因转移技术已经在小鼠[22-23]、牛[24-25]、猪[23]、鸡[26-27]、鱼类等生物中得到一定改良和应用,2014年Xin等[28]发现海洋鱼类精子由于精浆中存在DNA酶,摄取外源DNA能力较弱,可通过除去精浆并用脂质体包装DNA的方法实现精子介导基因转移。精子介导法有操作简便、成本低廉的优点,通过精子介导CRISPR/Cas9到节肢动物的受精卵或胚胎也作为引入CRISPR/Cas9的可行性方法受到研究者的关注,但在节肢动物中尚未报道。
1.3.4 纳米颗粒介导法。
用纳米颗粒包装CRISPR/Cas9系统进入细胞可以增大体内细胞对CRISPR/Cas9系统的内化率,克服离体基因编辑的限制,为内容物分子提供屏障以防止降解,还可通过设计纳米颗粒的结构和成分,灵活实现靶向特定的组织。CRISPR/Cas9进入细胞的3种方式产生的复合物与质粒均可以被纳米颗粒包装进入细胞[29-31]。对于通过纳米颗粒包装CRISPR/Cas9系统进入细胞来说,以Cas9 mRNA和sgRNA混合物的方式进行包装优于Cas9-sgRNA质粒包装,原因在于质粒DNA中包含了大量非编码序列,长度较大,使得包装难度高[32]。此外,mRNA可以直接在细胞质中进行翻译,不需整合到宿主基因组中,没有插入诱变的风险,脱靶率较低,质粒则需要进入细胞核中完整进行转录和翻译的过程。当前各种纳米颗粒的应用有很多,例如两性离子氨基脂质(zwitterionic amino lipids,ZALs)纳米颗粒[33]、外泌体-脂质体杂合纳米颗粒[34]、脂质纳米颗粒[35]和阳离子类脂质纳米颗粒[36-37]等都实现了递送CRISPR/Cas9进入细胞并实现基因编辑。但由于纳米颗粒传导法前期制备方法复杂、成本较高,因而在一定程度上限制了其在基因编辑操作中的广泛应用。