毕赤酵母对甲醇胁迫的响应及其对蛋白表达的影响
作者: 韩铭海 王未鲜 付相荣 彭意 康丽娟 龚勋 许存宾
摘要 醇氧化酶AOX1的启动子(PAOX1)被广泛应用于毕赤酵母表达外源蛋白,甲醇诱导可以使其表现出高效的转录活性,但甲醇诱导又可使毕赤酵母细胞产生过多的活性氧物质(ROS),诱发氧化胁迫响应和其他不利的生化生理响应,从而抑制蛋白的高效表达。综述了毕赤酵母对甲醇胁迫的响应及缓解甲醇胁迫的技术,并提出了UPR与甲醇胁迫响应交互作用机制的研究路线。
关键词 毕赤酵母;甲醇胁迫响应;氧化胁迫响应;非折叠蛋白响应
中图分类号 Q78 文献标识码 A
文章编号 0517-6611(2022)16-0001-04
doi:10.3969/j.issn.0517-6611.2022.16.001
开放科学(资源服务)标识码(OSID):
Research Progress on the Effect of Methanol Stress Response on Production of Heterologous Proteins in Pichia pastoris
HAN Ming-hai,WANG Wei-xian,FU Xiang-rong et al
(College of Food and Pharmaceutical Engineering,Guizhou Institute of Technology,Guiyang,Guizhou 550000)
Abstract The promoter of alcohol oxidase AOX1 (PAOX1) is widely used in Pichia pastoris to express heterologous proteins.Methanol induction will lead to its high transcriptional activity.However,the induction with methanol gives rise to the abundance of reactive oxygen species (ROS) in Pichia pastoris cells,resulting in oxidative stress response and other adverse biochemical and physiological responses.Accordingly,high-efficiency expression of proteins would be inhibited.In this paper,the response of Pichia pastoris cells to methanol stress and the technique of alleviating methanol stress are reviewed.The authors look forward to the technical route of studying the mechanism of the interaction between UPR and methanol stress response,and the strategy of stimulating UPR to alleviate methanol stress is also raised.
Key words Pichia pastoris;Methanol stress response;Oxidative stress response;UPR
毕赤酵母(Pichia pastoris,最新系统命名为Komagataella phaffii)是当今科研和商业化生产重组蛋白最常用、最受欢迎的表达平台之一。毕赤酵母具有良好的生物安全性、易高密度培养、高转录活性的启动子、较强的蛋白合成分泌能力、胞外产物易分离纯化及与更高级真核细胞类似的翻译后修饰等诸多优点[1-2]。毕赤酵母相比酿酒酵母(Saccharomyces cerevisiae)而言,其表达分泌蛋白的能力更卓越[1-2]。蛋白高水平表达技术和策略一直是该领域的研究热点,当前,提高蛋白表达的主要技术有[3-4]:一是优化发酵条件,包括培养基优化、诱导策略优化、高密度培养等手段;二是优化分子生物学技术,包括密码子优化、选用强启动子和高效信号肽、增加基因拷贝数等;三是改造蛋白折叠、修饰和分泌途径等。
在发酵过程中,毕赤酵母细胞会受到环境中的各种胁迫,例如代谢产物、不适温度、pH和离子强度等胁迫,这些胁迫可能会显著影响细胞的生理特性,包括蛋白合成和分泌表达能力,该问题已经受到越来越多的科研工作者的关注。毕赤酵母是甲基营养型酵母,能以甲醇作为唯一碳源和能源,同时,甲醇也是诱导毕赤酵母外源蛋白表达最常用的诱导物,但甲醇本身对宿主也会产生胁迫效应。因此,若要获得甲醇诱导外源蛋白的高水平表达,甲醇胁迫是需要关注的一个问题。
1 PAOX1的甲醇诱导
1.1 PAOX1的活性 毕赤酵母外源蛋白表达的启动子主要有醇氧化酶I(AOX1)、磷酸甘油醛脱氢酶(GAP)、醇脱氢酶(ADH3)、甲醛脱氢酶(FLD1)等基因的启动子,其中,AOX1的启动子(PAOX1)最常使用[5]。醇氧化酶包括AOX1和AOX2两种酶蛋白,毕赤酵母细胞85%的醇氧化酶活力来自AOX1,该酶蛋白催化甲醇转化为甲醛,是甲醇代谢途径中的关键酶;醇氧化酶与氧的亲和力较弱,因此,细胞需要合成大量的此酶蛋白来完成相应的催化功能[6]。在细胞利用甲醇生长过程中,PAOX1表现出极强的转录活性,AOX1占到了胞内蛋白的30%[5-6]。
1.2 诱导PAOX1的甲醇浓度 PAOX1在较低浓度甲醇条件下就可以被高效地诱导。例如,在0.5%浓度下,甲醇诱导毕赤酵母表达重组人豆荚蛋白(legumain)[7]、胰岛素前体(insulin precursor)[8]、耐热葡聚糖酶[9]和单链Fv抗体片段(single-chain Fv antibody fragment)[10]均能获得理想的表达水平。Vanz等[11]报道,诱导表达乙肝表面抗原(hepatitis B surface antigen)的甲醇浓度为0.6%。而Ohsawa等[12]报道,0.1%的甲醇浓度就能诱导PAOX1表现出极强的转录活性。
虽然大部分文献报道甲醇诱导外源蛋白表达最适浓度范围为0.5%~2.0%[1],但是也有学者报道,转录组学研究结果表明稍高浓度(10 g/L)的甲醇反而会抑制PAOX1的转录活性[12]。另外,除了PAOX1以外,毕赤酵母利用甲醇诱导的启动子还有PDAS、PFLD1、PPEX8等[5],利用这些启动子表达外源蛋白时,甲醇胁迫的影响同样是必须要关注的问题。
2 甲醇胁迫响应
甲醇不仅作用于毕赤酵母的甲醇代谢途径,而且对整个细胞都有影响[11,13]。Vanz等[11]报道在分批培养模式下,毕赤酵母对较低浓度的甲醇(6 g/L)即可产生胁迫响应。基于转录组学研究结果,毕赤酵母甲醇胁迫响应影响多个代谢途径和细胞生理特性[11,13]。涂庭勇等[14]也报道了采用“低甲醇浓度-高溶解氧浓度”策略诱导毕赤酵母能获得更高目标蛋白表达水平。由此,可以推测甲醇诱导外源蛋白表达的最佳浓度是甲醇诱导作用与甲醇胁迫效应两者平衡的结果。
2.1 上调甲醇代谢途径 毕赤酵母中,甲醇在过氧化酶体中被醇氧化酶(AOX1和AOX2)转化为甲醛和过氧化氢。甲醛在细胞质中被甲醛脱氢酶(FLD1)催化为甲酸,接着被甲酸脱氢酶转化为二氧化碳和水,并释放能量;或是甲醛在二羟丙酮合成酶(DAS1)作用下与5-磷酸木酮糖合成3-磷酸甘油醛和二羟丙酮,从而进入细胞同化代谢途径。在甲醇诱导阶段,70%~80%的甲醇进入了异代谢途径产生NADH和二氧化碳[11]。毕赤酵母甲醇诱导阶段甲醇代谢的关键酶类AOX1、甲酸脱氢酶(FDH1)、FLD1、甲酰谷胱甘肽水解酶(FGH1)和DAS1表达量上调[11,13],说明细胞甲醇代谢途径更活跃。
2.2 影响糖酵解途径、戊糖磷酸途径和TCA代谢途径
在甲醇诱导阶段,碳代谢主要集中于甲醇的代谢[15]。TPI1(磷酸甘油醛异构酶,引导甘油进入碳中心代谢途径)显著下调,糖酵解途径和戊糖磷酸途径酶类普遍下调;除了苹果酸脱氢酶(MDH1)显著下调外,TCA循环中的酶类变化不显著[11]。Li等[15]研究发现,在甲醇诱导阶段,毕赤酵母能量代谢途径普遍下调。
2.3 诱发氧化胁迫响应(oxidative stress response) 毕赤酵母需要提高过氧化物酶水平来抵御甲醇的毒害作用[16]。在甲醇代谢途径中,甲醇被AOX1和AOX2催化形成毒性更强的甲醛、H2O2和其他过氧化物及活性氧物质(ROS),导致细胞遭受氧化胁迫(oxidative stress),细胞进而上调一些过氧化物酶类,比如CTA1(去除H2O2)和PMP20(去除烷基氢过氧化物,维护过氧化酶体膜功能),来抵御氧化胁迫[11,13,15]。同时,氧化胁迫响应不仅提高毕赤酵母抵御氧化胁迫能力,也能增强其抗热应激能力[17-18]。
2.4 激活MAPK信号通路 一些特定的信号通路常常介导细胞对恶劣环境的应答,而这些信号通路大多数情况下与MAPK信号通路关联。毕赤酵母的甲醇诱导上调了MAPK信号通路相关基因的表达。MAPK信号通路与毕赤酵母细胞维持细胞壁的功能相关[19],与细胞生理生化和相关基因调节都有关系[13],影响细胞生长率;上调MAPK信号通路可有效地提高细胞抵御甲醇胁迫的能力,确保细胞的生存[13]。另外,对于外源蛋白表达来说,MAPK信号通路也影响PAOX1的活性[13]。
2.5 诱发细胞UPR 内质网对新生肽的合成及折叠效率对蛋白的高效表达至关重要[20-22]。在内质网中,新生肽经历极为复杂的折叠加工机制,而细胞严谨的质量控制机制确保了多肽的正确折叠。只有正确折叠的蛋白才能被细胞输送至高尔基体,接受进一步的修饰加工进而完成分泌表达,而非正确折叠多肽被内质网识别并进入ERAD途径,最终在细胞质中被蛋白酶体彻底消化。大量非正确折叠的多肽在内质网中集聚产生内质网胁迫(ER stress),上调细胞宿主未折叠蛋白响应(UPR)通路,以消除大量非正确折叠多肽对细胞的毒害作用[20-22]。
UPR通路虽然与内质网胁迫关系最为密切,但是其他胁迫,比如高温、高渗透压、有机溶剂、DTT、重金属、病毒、营养物饥饿等[20]都会激发此通路。UPR下游靶基因功能涵盖大部分新生肽合成、折叠、翻译后修饰、运输途径和ERAD途径等,对细胞生理特性影响也十分广泛,有些效应并不与蛋白表达有直接的关联。
有些报道虽然表明甲醇胁迫对UPR通路和ERAD途径的影响不显著[13]。但Vanz等[11]报道甲醇胁迫响应与毕赤酵母UPR通路和ERAD途径相关:PDI与SSC1蛋白(UPR通路关键蛋白)和CLPB与CDC48蛋白(ERAD途径功能蛋白)在甲醇诱导阶段上调了。而Li等[15]研究发现甲醇诱导阶段分子伴侣Hsp40上调了,表明激发了UPR;而ERAD途径是下调的。
2.6 诱发细胞自噬
在甲醇诱导阶段,细胞出现显著的自噬现象[11]:ATG1和其他与自噬相关的蛋白显著上调,而负调控细胞自噬途径的功能蛋白Tap42、Vps8、LCB1/2和Vps39则下调;天门冬酰胺蛋白酶(APR1)下调,标志着细胞液泡降解;与蛋白酶体相关基因表达上调[13];通过电镜观察到液泡由规则球形改变为内陷不规则形态[11,13];过氧化物酶体也呈现破坏并降解的现象[11,13],这也与细胞自噬有关。
3 甲醇胁迫响应与UPR通路的交互作用
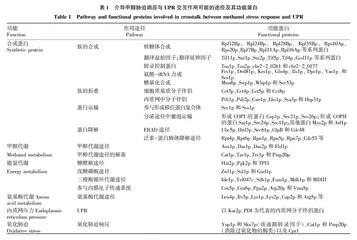
甲醇胁迫响应与UPR通路对毕赤酵母细胞的多个途径及生理特性都产生广泛的影响,两者之间存在交互作用的可能性。首先,甲醇胁迫导致上调细胞多个功能迥异的通路,包括上调与内质网应激机制相关的UPR通路,以缓解甲醇对细胞的胁迫,例如,甲醇诱导毕赤酵母表达一些外源蛋白上调了内质网中的分子伴侣蛋白,表明激发了UPR通路[11,13,23];其次,UPR与甲醇胁迫响应作用于细胞若干相同的途径,比如两者都作用于糖酵解途径、戊糖磷酸途径和ERAD途径,同样影响蛋白合成、运输和分泌途径等[11,13,23-24];最后,甲醇胁迫也会同时诱发UPR通路和氧化胁迫响应[11],而且在毕赤酵母中激发UPR和缓解氧化胁迫这两种策略都可以促进外源蛋白表达[18,25]。在酿酒酵母中已证实了UPR与氧化胁迫响应通路之间存在密切关联性[26-28],在毕赤酵母中可能也会存在类似的情况。因此,毕赤酵母UPR通路与甲醇胁迫响应存在交互作用的可能性,表1列举了介导甲醇胁迫响应与UPR交互作用可能的途径及其功能蛋白[11,24]。