MAPK信号通路调控植物响应非生物胁迫的研究进展
作者: 刘晨 曹小汉 殷丹丹 杨婧 张宁宁 任莉萍
摘要 丝裂原活化蛋白激酶(MAPK)级联信号通路是真核生物中广泛存在的信号转导途径。非生物胁迫是植物面临的首要挑战,随着极端气候的频发和环境污染问题的加剧,开展植物MAPK级联信号通路在非生物胁迫下的机理研究迫在眉睫。对近年模式植物拟南芥,主要农作物水稻、玉米和小麦等,以及重要园艺作物中MAPK信号通路响应干旱、盐胁迫、极端温度及营养匮乏等方面的研究进行了总结归纳,并对其进一步的研究工作进行了展望。结果表明,MAPK作用于植物响应非生物胁迫信号转导,并在植物抗逆过程中扮演重要角色。研究MAPK作用机制将对阐明植物抗逆分子网络,培育抗性品种和提高作物产量等方面具有重要意义。
关键词 植物;非生物胁迫;MAPK;信号通路
中图分类号 Q945.78 文献标识码 A 文章编号 0517-6611(2022)18-0009-08
doi:10.3969/j.issn.0517-6611.2022.18.003
开放科学(资源服务)标识码(OSID):
Research Progress of MAPK Signaling Pathway in Regulating Plants Response to Abiotic Stress
LIU Chen1, CAO Xiao-han2, YIN Dan-dan2 et al
(1.Nanjing Institute of Agricultural Sciences, Nanjing, Jiangsu 210046; 2. Biology and Food Engineering School, Fuyang Normal University, Fuyang, Anhui 236037)
Abstract Mitogen-activated protein kinase (MAPK) cascade signaling pathway is a widespread signal transduction pathway in eukaryotes. Abiotic stress is the primary challenge of plants. With the frequent occurrence of extreme climate and the aggravation of environmental pollution, it is extremely urgent to study the mechanism of MAPK cascade signaling pathway in plants under abiotic stress. In this paper, the response of MAPK signaling pathway to drought, salt stress, extreme temperature and nutrient deficiency in model plant arabidopsis, major crops (rice, maize and wheat) and important horticultural crops in recent years were summarized. The future researches of MAPK signaling pathway were prospected. The results show that MAPK signal transduction plays an important role in plant response and resistance to abiotic stress. The studies of the mechanism of MAPK will be of great significance to elucidate the molecular network of plant stress resistance, cultivate resistant varieties and improve crop yield.
Key words Plants;Abiotic stress;MAPK;Signaling pathway
相对于动物而言,植物在整个生命过程中通常都是无法移动的[1]。植物所生长的外界环境和其内部信号因子又会时时刻刻影响着植物的生长发育进程。其中非生物胁迫是严重影响植物生命活动的逆境之一,包括干旱胁迫、极端温度、营养匮乏、盐害、机械损伤以及土壤中有毒金属的毒害等。近年来极端天气越来越频繁地出现,非生物胁迫对农业生产的不利影响日益加剧,植物也面临着更加严峻的挑战[2]。植物为了在复杂且多变的外界环境作用下正常生长,进化出多个复杂而完善的信号转导和响应胁迫的调控网络[3]。因此,探索植物对外界非生物胁迫信号的感知和响应,对于培育优良抗性的新品种,进而降低水肥消耗,减少环境污染,促进农业绿色可持续发展具有重要的理论指导意义。
丝裂原活化蛋白激酶(MAPK)级联信号通路广泛存在于真菌、动物和植物中。近年来,科研工作者开展了大量有关MAPK信号通路的研究,发现植物在受到外源信号刺激后,利用多种机制将促分裂原活化蛋白的激酶激酶激酶(MAPKKK、MKKK/MEKK、MAP3K)激活,活化的MAPKKK通过磷酸化激活促分裂原活化的蛋白激酶激酶(MAPKK、MEK/MKK),进而激活促分裂原活化的蛋白激酶(MAPK、MPK)[4]。MAPK被磷酸化后可以进一步激活下游蛋白激酶和转录因子(如WRKY)等底物,这些激酶或转录因子可以通过调控相应功能基因的表达等方式,使植物细胞对外源信号产生特定的生理生化反应以应对环境变化[5]。根据蛋白催化区域氨基酸序列及结构的差异,将MAPKKK分为Raf、MEKK和ZIK 3类[6]。其中,Raf类的催化区域氨基酸序列为GTxx(W/Y)MAPE,MEKK类具有G(T/S)Px(W/Y/F)MAPEV保守结构域,ZIK类具有GTPEFMAPE(L/V)Y保守结构域。拟南芥是评价植物基因复杂性的蓝图,它含有80个MAPKKK基因,其中21个属于MEKK类,11个属于ZIK类,48个属于Raf类[7]。根据MAPKK蛋白结合、激活区的结构及氨基酸序列的差异,可将其分为A、B、C、D 4个亚型。MAPKK的氨基末端通常都有1个假定的MAPK对接结构域K/R-K/R-K/R-X1-6-L-X-L/V/I。A、C、D 3个亚型的MAPKK编码相对较短的蛋白质,而B亚型成员的羧基末端结构域与介导RanGDP核输入的NTF2(NUCLEAR TRANSPORT FACTOR 2)同源[8]。与MAPKKK相比,植物中的MAPKK基因数量较少。在拟南芥基因组中有10个MAPKK,其中有3个A亚型(MKK1、MKK2和MKK6),1个B亚型(MKK3),2个C亚型(MKK4和MKK5),4个D亚型(MKK7、MKK8、MKK9和MKK10)[9]。MAPK是整个MAPK级联系统的最下游的蛋白激酶。植物中的MAPK含有11个保守的亚结构域,在第七、八结构域间有1个三肽模块,即TxY基序,是被MAPKK磷酸化的位点。TxY基序位于蛋白质三维结构的T环结构(T-loop)上,它也是ATP的结合位点[8]。MAPK的激活是通过上游的MAPKK同时磷酸化TXY基序中的苏氨酸(Thr)和酪氨酸(Tyr)残基[9]。根据被MAPKK磷酸化的保守TxY基序(x通常为Glu/Asp),植物中的MAPK蛋白分为TEY和TDY 2个亚家族。含有TEY基序的亚家族又分为A、B、C 3类,含有TDY基序的MAPK是植物所特有的,组成了D类[8]。
目前已有大量的研究表明,MAPK信号通路与植物响应非生物胁迫有着密不可分的关系,并在增强植物抗逆性方面起着至关重要的作用。该途径可以通过磷酸化将非生物胁迫信号逐级放大、传递,以此来参与植物生长发育及抗逆作用等生物学过程[10]。该研究综述了已知的植物MAPK级联途径响应非生物胁迫信号转导的最新研究进展。分别在干旱胁迫、极端温度、营养匮乏和盐害等方面进行概述。以期为进一步阐明MAPK级联信号通路的分子调控网络提供一些参考,并为将来选育抗性品种、提高作物产量和农业可持续发展奠定基础。
1 MAPKs响应干旱胁迫
干旱是影响植物生长发育的一个严重的非生物胁迫因素,其造成的作物产量损失甚至比所有病原体伤害加起来还要多[11]。此外,干旱胁迫还会导致渗透胁迫、氧化胁迫、破坏细胞组分,并引起代谢紊乱等次级影响[2]。研究植物如何抵御干旱胁迫,将为筛选优良的抗旱农作物品种和提高农作物产量方面提供帮助。
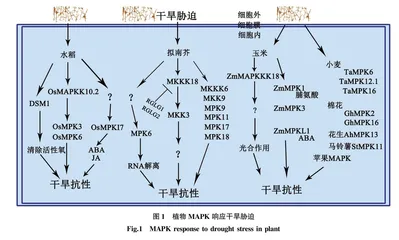
MAPK激酶级联信号通路在植物的抗旱机制中起着十分重要的作用(图1)[12]。有研究表明,对44个MAPK在不同非生物胁迫下的转录调控分析表明,拟南芥中的MPK9、MPK11、MPK17、MPK18、MKK9和MKKK6均受脱水胁迫诱导[13]。拟南芥MPKKK18能够激活MPKK3,MAPKKK18和MAPKK3超表达后耐旱性得到了增强,而泛素连接酶RGLG1和RGLG2抑制了这一过程,使耐旱性降低[14]。MPK6通过调节RNA解离活性来提高植物对脱水的耐受性[15]。水稻MAPKK10.2能够通过磷酸化激活MAPK6和MAPK3提高水稻的耐旱性[16]。OsMPK17蛋白质在干旱胁迫下诱导表达,且推测其功能受脱落酸(ABA)和茉莉酸甲酯(JA)等激素调控,超表达OsMPK17能够显著提升水稻在干旱条件下的生长势[17]。水稻中的DSM1(类似Raf的MAPKKK蛋白)作为活性氧的潜在清除剂,可以提高植物对水分胁迫下脱水的耐力,而突变体dsm1和RNA干扰植物对干旱胁迫更为敏感[18]。ZmMAPKKK18在玉米苗期抗旱过程中起重要作用,表达蛋白定位于叶绿体中,且在干旱胁迫下特异性调节光合作用的光反应阶段、吲哚乙酸生物合成途径Ⅱ、嘌呤核苷酸从头合成途径、产氧光合作用及有氧呼吸I等途径[19]。玉米ZmMAPK1超表达促进转基因拟南芥植株对干旱胁迫的耐受力,可增加干旱条件下的脯氨酸含量,降低丙二醛含量[20]。ZmMPK3也是干旱反应基因,提高了玉米的抗旱性[21]。MAPK-like(MPKL)蛋白是一组含有MAPK标志TxY基序的激酶,与MAPK序列相似,ZmMPKL1可能通过改变ABA生物合成和分解代谢基因的转录,维持ABA稳态来正向调节玉米幼苗对干旱敏感性[22]。小麦TaMAPK6(上调)、TaMAPK12;1和TaMAPK16(下调)等3个成员能够应答干旱胁迫[23]。棉花MAPK蛋白激酶GhMPK2在干旱条件下可以减少水分丧失、调节渗透压[24]。属于MAPK中D亚型的棉花GhMPK16也是干旱反应基因。过表达GhMPK16,在进行干旱处理后,植物体内H2O2含量升高,耐旱性则减弱[25]。花生AhMAPK13基因受干旱等胁迫的诱导上调表达[26]。在干旱条件下,马铃薯StMAPK11明显上调,并通过提高抗氧化活性(SOD、CAT和POD活性,以及过氧化氢、脯氨酸和MDA含量变化)来增强马铃薯植株的抗旱性[27]。此外,MAPK级联反应还参与调控植物和微生物互作从而增强对非生物胁迫的响应,干旱胁迫下,苹果植株和丛枝状菌根中的MAPK均表达上调,推测菌根寄生利用MAPK信号与苹果植株的相互作用,从而增强苹果的耐旱性[28]。
2 MAPKs响应盐胁迫
土壤中的盐度会影响到植物的光合作用、蛋白质代谢和脂质的代谢等活动,进而影响到植物的生长发育。高盐浓度还会进一步导致次级胁迫伤害,如离子毒性(主要是Na+)、高渗胁迫以及氧化损伤等[2]。相应地,植物也会进化出多种机制来抵御盐胁迫。目前已经有许多研究发现MAPK能介导植物的抗盐胁迫(图2)。
拟南芥中发现了一个MAPK信号通路,是由MKKK1、MKK2和MPK4/MPK6组成。过表达MKK2的植株会表现出更强的耐盐性[29]。拟南芥mkkk20突变体对高盐胁迫敏感,在脱水条件下的水分流失率高于野生型(WT)植株,且活性氧(ROS)积累增加。相反,过表达MKKK20的转基因植株对盐胁迫表现出耐受性[30]。MPK6激活并与MYB41相互作用,使拟南芥的耐盐性增加[31]。此外,在盐胁迫下,磷脂酸(PA)对MPK6有调节作用,这表明MPK6和PA可能是通过同一信号通路发挥作用的[32]。盐胁迫条件下MKK3-MPK6级联激活拟南芥MYC2(bHLH类转录因子),通过调节脯氨酸生物合成从而调控盐胁迫抗性[33]。拟南芥愈伤组织受到盐胁迫处理下,MKK9-MAPK3/MAPK6通路在增加替代呼吸中起着关键作用[34]。AtMPK9可被盐胁迫激活,且在体外可以独立于级联而进行自磷酸化作用[35]。在水稻中OsMPK44、OsMPK5和OsMPK4也被证明与抗盐胁迫有关。过表达OsMPK5和OsMPK44的水稻转基因株系会表现出更高的耐盐性[36]。在玉米中,ZmSIMK1的表达也受到盐胁迫调节,过表达此激酶并用盐胁迫处理,RD29A和P5CS1基因表达量上升,而且玉米耐盐性也得到了提高[37]。高盐处理下,小麦TaMPK4、TaMPK6和TaMPK17表达水平逐渐升高,在正常生长浓度下,它们的表达水平降低。而高盐处理下TaMPK1和TaMPK16表达水平下降,随着恢复处理的增加,它们的表达水平逐渐增加[22]。桑树MnMAPK1也受盐胁迫诱导,且其拟南芥过表达株系具有更强的耐盐性[38]。
3 MAPKs响应极端温度胁迫
温度对植物的生长发育有很大的影响,植物能够在适合的温度范围内正常生长,一旦超出范围,植物可能就会受到极端温度的胁迫。低温会限制种子萌发、植物生长和新陈代谢。而高温会引起热应激并加速呼吸作用和光合速率,从而使活性氧产量过剩,影响植物生长[39]。近年来,极端天气气候事件频发,这将对作物带来更大的挑战[2]。植物如何抵御极端温度胁迫,也一直是科研工作者们关注的焦点。