裂解多糖单加氧酶在植物病害中的研究进展
作者: 陈进银 张莉 郭秀娜
摘要 植物病原真菌在侵染植物的过程中会分泌细胞壁降解酶来破坏植物细胞壁,以达到成功入侵寄主的目的。研究表明,病原菌分泌的细胞壁降解酶中的纤维素酶、果胶酶、木聚糖酶、几丁质酶、酯酶均参与了植物细胞壁的降解和病原菌致病的过程,但是对于细胞壁降解酶中的裂解多糖单加氧酶在病原菌侵染植物过程中的功能研究较少。根据近年来辅助活性酶类以及属于辅助活性酶类中的裂解多糖单加氧酶的研究进展,综述了辅助活性酶的分布、种类、功能以及裂解多糖单加氧酶植物病害中的作用研究,旨在为辅助活性酶类在植物病害中的功能研究和病害防控提供参考和依据。
关键词 裂解多糖单加氧酶;细胞壁降解酶;辅助活性酶;真菌病害
中图分类号 Q945.8 文献标识码 A 文章编号 0517-6611(2024)03-0011-04
doi:10.3969/j.issn.0517-6611.2024.03.003
Research Progress of Lytic Polysaccharide Monooxygenase in Plant Diseases
Abstract During the process of infecting plants,phytopathogenic fungi secrete cell wall degrading enzymes to destroy plant cell walls in order to successfully invade the host.Studies have shown that cellulase,pectinase,xylanase,chitinase,and esterase in the cell wall degrading enzymes secreted by pathogenic bacteria are involved in the process of plant cell wall degradation and pathogenic bacteria.However,there is relatively little research on the function of the lytic polysaccharide monooxygenase in the cell wall degrading enzymes in the process of pathogen invasion of plants.Based on the recent research progress of auxiliary active enzymes and cleavage polysaccharide monooxygenases belonging to the auxiliary active enzymes,this article summarizes the distribution,types,functions of auxiliary active enzymes and the role of cleavage polysaccharide monooxygenases in plant diseases.The research aims to provide reference and basis for the function research and disease prevention and control of auxiliary active enzymes in plant diseases.
Key words Lytic polysaccharide monooxygenase;Cell wall degrading enzyme;Auxiliary active enzyme;Fungal disease
植物在正常生长发育过程中会遇到各种各样的不利因素,如极端天气、干旱、涝害、植物病害等,最终影响植物的产量和质量,其中在生物因素中病原菌对植物生长发育影响较大。植物与病原真菌为了各自的生存,在数千年的相互对抗中进化出了各种抵抗和入侵对方的策略。植物对抗病原菌的第一道屏障是植物细胞壁,主要由多糖物质中的纤维素和果胶组成,而病原菌为了成功入侵寄主植物获取自身所需营养,进化出了一系列策略来破坏植物细胞壁,其中病原菌分泌细胞壁降解酶来降解植物细胞壁是最主要的一个策略。研究发现,碳水化合物活性酶数据库(carbohydrate-active enzymes database,CAZy,www.cazy.org)中的糖苷水解酶(glycoside hydrolases,GHs)、多糖裂解酶(polysaccharide lyases,PLs)、碳水化合物酯酶(carbohydrate esterases,CEs)中的成员均参与了植物细胞壁的降解过程和病原菌的致病过程[1-6]。而CAZy中还有一类主要的酶类——辅助活性酶(auxiliary activities,AAs),其可以氧化裂解顽固多糖,但是对于其在病原菌致病过程中的功能研究较少。
目前,辅助活性酶类主要分为木质素分解酶家族和裂解多糖单加氧酶家族(lytic polysaccharide mono-oxygenases,LPMO),其中在病原菌致病过程中仅有LPMO的研究成果。LPMO是一种铜离子依赖的氧化酶,其可以通过氧化和裂解的方式解聚顽固多糖中的糖苷键,对生物质转化具有积极意义。
1 辅助活性酶的研究进展
AAs原来归类于糖苷水解酶(61家族糖苷水解酶——GH61和33家族碳水化合物结合模块——CBM33),但是研究发现其并不能通过水解的方式降解纤维素,而是通过氧化的方式裂解纤维素形成纤维寡糖,后来将其重新划分归类于辅助活性酶家族[7]。
AAs本身不具有纤维素水解酶的活性,但是近年来由于其可以显著提高纤维素水解酶的活性而备受关注[8-9]。AAs在核酸序列和蛋白结构上具有特殊性,AAs各家族成员的核酸序列保守性相对较低,但是其蛋白结构中的酶活性中心相对保守,一般都2个组氨酸和1个酪氨酸形成的三N配体“组氨酸支架”和1个二价金属离子构成,并且其中1个组氨酸必须为成熟蛋白N端的第1个氨基酸,这对于AAs的酶活性是必须的[10-12]。
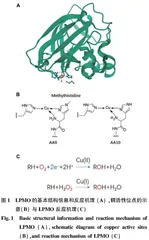
目前对于AAs的研究主要集中于蛋白结构、底物的种类、电子供体、氧化方式、共底物、酶动力学等方面(图1)[13-19]。图1A为 LPMO的基本结构信息和反应机理,来自解淀粉芽孢杆菌的 AA10 LPMO BaAA10 的蛋白结构,其中活性位点铜显示为球体,活性位点残基显示为棒状; 图1B为在AA9和AA10结构中观察到的铜活性位点示意; 图1C为 LPMO 反应机理,箭头上方指示的 Cu(II)/Cu(I)是指活性位点中的铜离子及其在催化循环开始之前的氧化态[20] 。
2 真菌中辅助活性酶的分布、种类和功能
2.1 种类与功能
在CAZy数据库中将AAs分成了17个家族,AA1~AA17。AAs类目前分为9个木质素分解酶家族(AA1~AA8、AA12)和8个裂解多糖单加氧酶家族(AA9~AA11、AA13~AA17)。木质素分解酶可能不作用于碳水化合物,但是由于木质素与植物细胞壁中的碳水化合物密切相关,而且木质素分解酶确实与经典的多糖解聚酶共同作用,所以CAZy 数据库根据氨基酸序列的相似性对木质素分解酶家族在家族和亚家族中进行了分类,意图反映蛋白质结构特征并促进基因组注释。目前对于LPMO的研究比较深入。LPMO可以氧化裂解糖苷键形成寡糖,但主要作用底物以及氧化切割底物的方式有所不同。AA9家族成员可以通过碳C1和/或C4和C6的氧化方式裂解纤维素链形成纤维寡糖;AA10家族成员可以通过碳C1和/或C4氧化裂解几丁质;AA11家族成员可以通过碳C1氧化裂解几丁质;AA13家族成员可以通过碳C1氧化裂解淀粉;AA14家族成员可以通过碳C1氧化裂解木聚糖;AA15家族成员可以通过碳C1和/或C4氧化裂解纤维素和几丁质;AA15家族成员可以通过碳C1氧化裂解纤维素;AA17家族成员可以通过碳C4氧化裂解同型半乳糖醛酸。
2.2 分布
AAs家族成员在可以降解纤维素的真菌与细菌的降解酶系统以及病毒中广泛存在[21-23]。其中,真菌中的AAs主要存在于担子菌门和子囊菌门中[24-26]。此外不同营养类型的植物病原真菌所拥有的数目也差异巨大,例如,活体营养型病原物小麦白粉菌Blumeria graminis的基因组[27]中AA9家族基因的数量明显少于其他真菌,其中包含1个AA9家族基因[28];死体营养型植物病原真菌如葡萄孢属Botrytis cinerea有10个AA9家族基因,立枯丝核菌Rhizoctonia solani有34个AA9家族基因。现在,大量来源于致病菌的PMO基因已被编入CAZy数据库(表1)和NCBI(https:∥www.ncbi.nlm.nih.gov/)数据库中,但是其功能大部分都未被鉴定。表1以AAs中第9家族(AA9)成员为例,展示部分植物病原菌中AA9家族基因的数量。
2.3 AA9 家族 LPMO 的底物特异性
AAs类可以解聚顽固多糖,目前不同AA家族成员已被证明具有多种底物活性,包括氧化裂解结晶纤维素、半纤维素、几丁质、木聚糖、木葡聚糖、淀粉等[13,18,29]。目前病原真菌中的多个LPMO也被证明有多重底物活性(表2),然而这些成分同时也是植物细胞壁的组成成分。这些结果暗示在病原菌侵染植物寄生的过程中LPMO是不可或缺的,其可以破坏植物细胞壁而帮助病原菌的成功入侵。
3 裂解多糖单加氧酶在病原菌与植物互作过程中的功能研究
LPMO最先在真菌和卵菌中被发现,其中,AA9是迄今为止数量最多的LPMO家族[34]。在尖孢镰刀菌(Fusarium oxysporum)侵染豆科植物根部时,几个编码AA9的基因均上调表达[35],同样葡萄孢属真菌侵染葡萄果实过程中,编码AA9的相关基因(BC1G_07653andBC1G_07658)也明显上调[36],这说明AA9编码基因可以在侵染植物时于多个组织和器官中发挥作用。
稻瘟病菌(Magnaporthe oryzae)附着胞成功定植的关键因素之一是其对表面信号的响应。研究发现,稻瘟病菌中的Aa91(MoAa91)是辅助活性酶第9家族的一个裂解多糖单加氧酶,MoAa91是一种受RGS和RGS样蛋白调控的新型信号分子。MoAa91可以与植物体内的免疫受体几丁质诱导结合蛋白前体(CEBiP)竞争几丁质结合,进而抑制由几丁质诱导的植物免疫应答进程,并且发现MoAA91受转录因子MoMsn2的调控,其破坏可以导致人工诱导表面附着胞的形成缺陷和病原菌的完全毒力缺陷[37]。
AA9 家族的 LPMO 在生物界广泛保守,被植物病原体部署以裂解纤维素聚合物。作为回应,植物进化出复杂的机制来感知细胞壁损伤,从而自我触发损伤触发免疫反应。非致病性病原真菌嗜热硫杆菌(Thermothielavioides terrestris)LPMO家族的TtAA9E降解纤维素产生的C1和C4氧化产物AA9_COS在拟南芥遭受死体营养型病原真菌灰霉病菌(Botrytis cinerea)侵染时可以作为DAMP(Damage-associated molecular pattern)保护植物免受侵染,造成胼胝质积累、病斑大小减小以及植物激素水平提高,引起拟南芥体内转录重编辑,最终导致对坏死性真菌灰葡萄孢的抵抗力增加,表明 LPMO 可能在植物病原体相互作用中发挥关键作用[38]。