NAC转录因子:参与植物诸多生命过程的调控因子
作者: 张兆冉 王利娜 朱博艺 李朝炜
摘要 NAC(NAM、ATAF1/2、CUC2)转录因子家族是植物中最大的蛋白家族之一,在多种植物中均发现其家族成员。NAC转录因子(NAC transcription factors,NAC TFs)一般由保守的N端特殊的DNA结合域以及多样的C端调控结构域组成,其数量十分庞大且功能多种多样,主要通过调控靶基因的表达水平参与调控诸多生命过程,进而调控植物的次生代谢产物合成、生长发育和应激反应,探究其生物学功能及分子调控机制,对改良植物农艺性状具有重要意义。系统综述了NAC TFs的研究现状、结构特点、作用模式和进化历程,及其对植物生命活动的调控包括次生生长、生物合成、生物/非生物胁迫响应、器官衰老等,旨在为改良植物农艺性状提供理论基础。
关键词 NAC;功能;生长;胁迫;激素响应
中图分类号 Q943.2 文献标识码 A 文章编号 0517-6611(2024)15-0022-08
doi:10.3969/j.issn.0517-6611.2024.15.005
开放科学(资源服务)标识码(OSID):
NAC Transcription Factors: Regulatory Factors Involved in Many Life Processes of Plants
ZHANG Zhao-ran,WANG Li-na,ZHU Bo-yi et al
(College of Food Science and Biology,Hebei University of Science and Technology,Shijiazhuang,Hebei 050000)
Abstract The NAC(NAM,ATAF1/2,CUC2)protein family is one of the largest protein families in plants,and its members are detected in many plants.The NAC protein family is involved in the regulation of many plant life processes as a transcriptional regulator.NAC transcription factors (NAC TFs) generally consists of a special N-terminal DNA binding domain and a variety of C-terminal transcriptional activation / inhibition regions.The number of NAC TFs is very large and their functions are diverse,which plays an important role in regulating plant life activities.Therefore,it is of great significance to explore their biological functions and molecular regulatory networks for improving plant agronomic traits.In recent years,the research on NAC TFs has been hot.This paper systematically reviewed the research status,structural characteristics,action mode and evolution process of NAC TFs,as well as its regulation of specific plant life activities including secondary growth,biosynthesis,biotic/abiotic stress,hormone response,organ senescence,etc.,and prospected its research trend.
Key words NAC;Function;Growth;Stress;Hormone response
在植物正常生命活动中基因的有序表达是维持生命进程的重要环节,而在基因表达中转录因子又是最基本的参与元件之一。NAC家族成员广泛分布在陆生植物中,其调控作用在多种植物中均有报道。1996年,Souer等[1]在矮牵牛中发现第1个NAC基因NAM(no apical meristem),其参与矮牵牛顶端分生组织的发育;而后在拟南芥中发现ATAF1/2(arabidopsis transcription activation factor 1/2)基因和CUC2(cup-shaped cotyledon 2)基因[2],前者编码重要的转录激活因子,后者的表达产物可影响植物叶片的发育,以上3个基因具有不同的生物学功能,但均有相似的N端结构域,NAC的命名即根据具有该结构域的这3个基因的首字母而来。在随后的研究中,NAC TFs在水稻(Oryza sativa)、小麦(Triticum aestivum)、白菜(Brassica rapa)、猕猴桃(Actinidia chinensii)、辣椒(Capsicum annuum)和花生(Triticum aestivum)[3]等多种植物中被发现。大量研究表明,NAC TFs参与植物诸多生命活动。
1 NAC TFs的结构特点
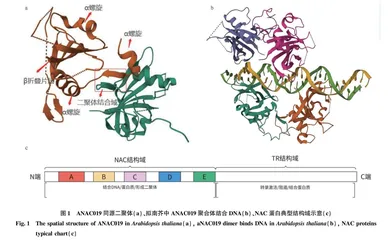
NAC蛋白家族一般由200~500个氨基酸组成,具有典型的空间结构,由几个α螺旋围绕中央一个反向平行的β折叠片层组成,中央β折叠直接与DNA结合,以拟南芥中的ANAC019的三维结构为例,3个α螺旋围绕1个β折叠,2个ANAC019通过N端二聚体结合域形成二聚体[4],见图1。NAC TFs的N端较为保守,由150~160个氨基酸组成[5],通常分为5个子域(A~E),与靶DNA的识别和结合十分重要。ACD 亚结构域保守性较高,且与NAC二聚体的形成以及DNA结合等过程相关;B、E亚结构域较其他3个子域保守性低,可能和NAC TFs的功能多样性有关。大部分的NAC蛋白的CD结构域中有核定位信号,引导NAC转录因子于细胞核中发挥作用。C端为转录调控区域,与N端相比其保守性较差,且具有多样性,该端往往富含一些出现频率很高的简单氨基酸,例如:丝氨酸、苏氨酸、谷氨酸等[6]。其可通过调控NAC转录因子与靶蛋白相互作用发挥功能。研究表明,C端的基序对于一个特定的NAC亚家族往往是保守的,而不同NAC亚家族的C端基序差异明显。部分NAC蛋白的C端也有跨膜结构,可与细胞质膜或内质网结合,称为NAC膜结合的转录因子(NAC membranebound transcription factors,NAC MTFs)。有些NAC蛋白的N端有一个特殊的锌指结构域(zinc finger domain),或有2个重复串联的N端结构域,甚至缺乏C端结构域[5],还有一些NAC TFs的启动子上含有多个响应激素信号和逆境胁迫的元件,表明NAC TFs受到激素影响的同时参与调控植物的逆境胁迫反应[7]。
2 NAC TFs的系统发育和分类
NAC基因家族成员几乎存在于所有陆地植物中,从“简单”苔藓植物到“复杂”被子植物,甚至存在于原始链生绿藻中。根据系统发育分析,NAC TFs的起源可能发生在距今约7.25亿~12.00亿年前的链状植物(Streptophytina)的多样进化时期,其结构域的中央β折叠与WRKY蛋白家族的DNA结合域有着同源的拓扑结构,因此推测其是原始真核生物的WRKY蛋白进化而来的[8-9]。链状植物包括淡水和陆生两大部分。在大约4.70亿年前,陆生苔藓植物出现,为适应陆生环境,NAC TFs家族出现了第一次扩张,主要是参与调控水分传导和提供机械强度的基因;第二次扩张发生于1.45亿年前的早白垩纪,与被子植物的进化同步[10]。
Pereira-Santana等[11]通过对24种陆地植物中3 187个NAC蛋白进行比较基因组分析,将这些蛋白分为6个主要类群(Ⅰ~Ⅵ组)。NAC Ⅰ组蛋白主要包括维管NAC结构域(vascular NAC domain,VND)亚组和次生壁相关NAC结构域蛋白(secondary wall-associated NAC domain protein,SND)亚组,主要参与最基本的生命活动,包括细胞壁组成、生物合成、水分传导和木质部发育等。NAC Ⅱ组中的蛋白主要参与植物的生长发育,例如茎尖分生组织(SAM)形成和调控子叶分离等。NAC Ⅲ组被称为跨膜基序(transmembrane Motif,TMM)组,TMM基序受膜内蛋白酶的水解切割调控,通常由生物和非生物胁迫信号介导。NAC Ⅳ组包括参与控制多个发育过程时序的蛋白,例如拟南芥开花时间调控基因LOV1[12]和拟南芥寿命主效调控基因ANAC042[13]等。NAC Ⅴ组包含多个参与应激反应和衰老过程的蛋白成员。NAC Ⅵ组包括多种含有被子植物特异性序列的NAC TFs。随着植物生存的需要,NAC TFs数量不断扩增且功能趋于多样化。
3 植物NAC家族的功能研究
NAC家族具有时间特异性,在植物不同发育时期和多种环境因素诱导下,激活特定基因的表达,发挥生物学功能。NAC蛋白家族数量众多且功能各异,在植物生长、激素调节、响应外界环境胁迫等基本生命活动中发挥作用。随着对植物基因组的深入研究,越来越多的NAC TFs被发现(表1)。不同NAC TFs在植物系统中的作用见图2。
3.1 参与植物的次生生长
在植物的生活史中,次生生长是一重要的生命活动,细胞壁次生加厚、木质化程度增加,帮助植物抵御机械损伤的同时控制水分散失,次生维管组织发育增强了植物的养分运输和机械支撑,尤其是一些木本乔木的次生生长,对人们的生产生活有重要意义。在NAC TFs中有很多成员参与次生生长过程,在拟南芥中NAC次生壁加厚促进因子(NAC secondary wall thickening promoting factor,NST)、SND、VND被发现共同调控拟南芥次生生长过程,NST1/2参与花药药室内壁的次生加厚,SND亚组中SND1参与维管组织中次生壁发育,并调控其他SNDs的表达,SND2/3参与纤维细胞次生壁发育,VND1~7被证实影响次生木质部发育以及导管的形成[36-37]。Zhong等[38]在拟南芥中鉴定出1个NAC基因ANAC099,其表达产物称为次生壁相关NAC结构域蛋白5(SND5),ANAC099在木质部导管中特异表达,正调控拟南芥次生壁合成,同时研究发现SND5及其同源物SND4(ANAC075的表达产物)在次生壁形成细胞特异表达,如果抑制其功能,植物的次生壁增厚明显被抑制,进一步研究发现SND2/3/4/5通过结合次生壁NAC结合元件(secondary wall NAC binding elements,SNBEs)调控靶基因表达,SND2/3与次生壁NAC主开关(secondary wall NAC master switch,SWNs)MYB46 / MYB83结合而后激活下游次生壁相关基因的表达,MYB46 / MYB83已经被证实能够诱导植物次生壁纤维素、木聚糖和木质素的积累。白桦BpNAC5参与调控次生壁的合成,BpNAC5过表达植株的最终株高高于野生型(WT),且其纤维素含量、木质素含量明显高于野生型,而半纤维素含量低于野生型,说明BpNAC5基因正向调控植物次生壁的生长以及组分的合成[25]。2021年Kim等[27]利用RNA测序(RNA sequencing,RNA-seq)和Pac Bio SMRT等测序技术对赤松维管组织进行特异性转录组分析,从62个NAC家族成员中鉴定了7个在木质部发育组织中高表达的PdeNAC基因,并证明了4个与VND(vascular-related NAC domain)、NST和SND4/ANAC075同源的PdeNAC基因(PdeNAC2、PdeNAC9、PdeNAC38、PdeNAC41)均可促进木质部导管细胞的发育。
3.2 调控植物根的生长
NAC家族在调控植物根的生长发育中起到关键作用。在拟南芥中E2启动子结合因子(E2 promoter-binding factors,E2Fs)是调控细胞分裂、DNA修复和细胞分化的重要转录因子[39]。E2启动子结合因子a(E2Fa)已被证明通过控制G1/S细胞周期转换来调节根的生长。拟南芥NAC1直接作用于E2Fa的启动子从而抑制E2Fa的表达,进而通过调控核内周期来控制根分生组织细胞分裂,促进根部生长。nac1突变体主根发育明显不良,根尖分生组织萎缩,分生组织细胞数量减少,而NAC1过表达株系主根显著长于WT,根尖分生组织增大,分生组织细胞数量增加,同时发现NAC1过表达株系中E2Fa表达下调[40]。OsNAC2在调控水稻根系和节间生长中起到重要作用,OsNAC2上调与生长素(IAA)失活相关基因(GH3.6和GH3.8)的表达,下调生长素(IAA)反应基因(OsARF25)和细胞分裂素(CTK)氧化酶基因(OsCKX4)的表达,从而增强细胞分裂素应答,但削弱根系生长中的生长素应答,负调控根系生长[41]。Xu[42]通过筛选拟南芥NAC蛋白亚家族Ⅲ,发现了核定位的受硝酸盐诱导的转录激活因子基因NAC056,NAC056可促进侧根生长并增强其在硝酸盐缺乏胁迫时的耐受性。进一步研究发现,NAC056调控NO3-同化所需基因的表达,直接靶向结合硝酸盐同化关键基因NIA1的启动子促进其表达,NIA1编码硝酸还原酶,同时正调控拟南芥侧根的发育。在高硝态氮环境中,NAC056促进了硝态氮同化,进而促进了根系生长。小麦TaNAC2-5A同样受硝酸盐诱导表达,染色质免疫共沉淀试验表明,TaNAC2-5A可以直接结合到编码硝酸盐转运蛋白和谷氨酰胺合成酶基因的启动子区域,在小麦中过表达TaNAC2-5A促进了根系生长和硝态氮内流速[43]。2019年Yang等[44]利用拟南芥过表达载体研究大豆NAC基因GmNAC109的功能,序列分析表明,GmNAC109与调控生物和非生物胁迫响应的ATAF1A高度同源。过表达GmNAC109可以提高非生物胁迫耐受性并促进侧根的形成。之前的研究发现,拟南芥中异位表达大豆GmNAC20,GmNAC20通过调控DREB/CBF-COR、生长素信号通路,提高转基因拟南芥的耐盐性和抗冻性,并诱导侧根的形成[45]。Yarra等[46]发现,在水稻中过表达GmNAC20,可显著提高非生物胁迫响应相关基因(OsDREB1A、OsDREB1B、OsDREB1C、OsICE1、OsTPP1、OsLEA3、OsP5CS、SNAC1)的表达水平,增强其耐盐性和耐寒性;同时也提高了生长素相关基因(OsIAA11、OsIAA13、OsAUX1)的表达,使转基因水稻侧根数量明显增加。