植物SPL转录因子研究进展
作者: 葛奇 席会鹏
摘要 SPL(squamosa promoter-binding protein-like)基因编码的绿色植物所特有的转录因子,参与植物形态建成、花发育、根发育等过程,调控植物不同发育时期的转变,维持植物育性,响应植物的外界胁迫,在植物的整个生长发育过程中发挥着重要的作用。对SPL转录因子的研究概况、结构特征、同源基因克隆情况、表达调控模式以及生物学功能进行了综述,以期为进一步进行SPL转录因子的相关研究奠定基础。
关键词 SPL;转录因子;基因克隆;表达调控;生物学功能
中图分类号 Q943.2 文献标识码 A 文章编号 0517-6611(2023)23-0025-05
doi:10.3969/j.issn.0517-6611.2023.23.006
Research Progress of SPL Transcription Factors in Plants
GE Qi1,XI Hui-peng2
(1.College of Landscape Gardening,Southwest Forestry University,Kunming,Yunnan 650224;2.Xishuangbanna Tropical Botanical Garden,Chinese Academy of Sciences,Mengla,Yunnan 666303)
Abstract SPL(squamosa promoter-binding protein-like) gene encodes a transcription factor unique to green plants,which participates in plant morphogenesis and flower development, root development and other processes, regulates the transformation of plants at different developmental stages,maintains plant fertility,responds to external stresses of plants,and plays an important role in the whole process of plant growth and development. The research situation, structural characteristics, homologous gene cloning, expression regulation pattern and biological functions of SPL transcription factors were reviewed in this paper,in order to lay the foundation for further research on SPL transcription factors.
Key words SPL;Transcription factor;Gene cloning;Expression regulation;Biological function
基金项目 国际合作课题(Y9HX111B02)。
作者简介 葛奇(1996—),女,云南保山人,硕士研究生,研究方向:风景园林植物资源及应用。*通信作者,高级工程师,硕士,从事植物资源保护及利用研究。
收稿日期 2022-11-07
SPL(squamosa promoter-binding protein-like)基因所编码的转录因子,是绿色植物特有的一类转录因子,称为SPL转录因子,广泛存在于单细胞绿藻、苔藓、裸子植物以及被子植物中。最早的2个SPL基因由Huijser 等[1]从金鱼草(Antirrhinum majus)花序中得到,因其具有能够识别并结合SQUAMOSA启动子的活性而被命名为SBP1和SBP2[2-3],即SQUAMOSA启动子结合蛋白(squamosa promoter binding protein,SBP)。其后又有多名研究人员在各植物中相继发现多个SPL转录因子,并证实该类转录因子在植物的形态建成、花器官发育及开花、果实发育和成熟、调控植物次生代谢、影响植物激素信号转导、参与外界胁迫应答等方面发挥着重要作用。
该研究在前人研究的基础上,搜集整理SPL转录因子近年来的研究成果,概括了当前的研究概况,阐述了SPL转录因子的结构特征,说明了SPL基因家族的克隆情况,分析了SPL转录因子的表达模式和生物学功能,并对其研究发展趋势进行了展望,以期为植物SPL转录因子的研究发展提供有价值的参考依据。
1 SPL转录因子研究概况
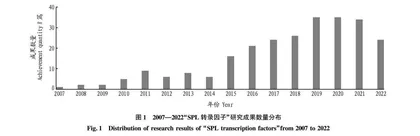
近年来,随着国内外对于SPL转录因子的研究逐渐深入,相关研究文献资源不断递增。对国内外研究成果进行简要分析,可以促进对SPL转录因子研究情况的全面了解,从而准确把握SPL转录因子相关的研究动向,促进SPL转录因子的研究发展。以中国知网(CNKI)为数据源,设定主题词为“SPL转录因子”,限定学科范围与植物有关,共检索到研究论文254篇,其中学术期刊53篇,学位论文196篇,国内会议5篇。相关报道显示,国内SPL转录因子的研究起始于2007年,并在近年快速发展(图1),内容大都集中在基因功能的研究方面,对于其上游调控基因及下游靶基因的研究相对较少。而国外早在1986年就开始了关于SPL转录因子特异性启动子的研究[4],且内容范围不仅仅局限于基因功能的研究,在基因的表达调控模式等方面的研究成果也十分丰硕。由此可见,国内对于SPL转录因子的研究还应进行更加深入全面地探讨,以准确详尽地掌握其基因表达调控的具体模式和内在的分子机理。
2 SPL转录因子的结构特征
SPL转录因子带有一个约由79个氨基酸残基组成的高度保守的DNA结合域,即SBP结构域(SQUAMOSA promoter-binding protein domain)[5]。SBP结构域是典型的锌指结构,由8个半胱氨酸(Cys)和组氨酸(His)残基组成,其中前4个氨基酸残基结合一个锌离子,后4个氨基酸残基结合另外一个锌离子[6]。目前发现的SBP蛋白中还存在一个位于该结构域C端的保守核定位信号。该信号能够与第2个锌指结构发生部分重叠,从而引导SBP蛋白进入植物细胞的细胞核,并进行表达[7]。
目前,在绿色植物中,被鉴定出来并根据基因结构差异分类SPL基因已有许多报道。在拟南芥(Arabidopsis thaliana)中,SPL基因家族共有17个成员。其中第一类成员含有10个及以上的外显子,所编码的蛋白质含有超过800个氨基酸残基,主要包括AtSPL1、AtSPL7、AtSPL12、AtSPL14、AtSPL16;第二类含有2~4个外显子,其SBP蛋白质不超过400个氨基酸,主要包括AtSPL2、AtSPL3、AtSPL4、AtSPL5、AtSPL6、AtSPL8、AtSPL9、AtSPL10、AtSPL11、AtSPL13、AtSPL15、AtSPL17[8]。在葡萄(Vitis vinifera)等植物中,陈文文等[9]根据miR156靶点的分布,将SPL基因家族分为9个主要分支,其中第一、二、三分支不包含miR156靶基因,第四、五、七、八、九分支其基因的CDS区内含有miR156靶点,第六分支其基因的3’UTR区含有miR156靶点,由此可见miR156位点在不同植物中具有较高的保守性。
3 SPL同源基因的克隆分析
已有研究证实SPL基因从简单的单细胞藻类植物到复杂的高等植物均有分布,如从衣藻(Chlamyydomonas reinhardtii)到小立碗藓(Physcomitrella patens)再到高等植物,其存在数量不一[3,10]。随着科技的进步,近年来已有大量SPLs基因被鉴定,如在拟南芥(Arabidopsis thaliana)中有17个,玉米(Zea mays)中有42个,水稻(Oryza sativa)中有19个,番木瓜(Carica papaya)中有10个,葡萄(Vitis vinifera)中有18个,菠萝[Ananas comosus (L.) Merr.]中有16个,海岛棉(Gossypium barbadense)和陆地棉 (G.hirsutum)中各有59 个,而人参(Panax ginseng C.A.Meyer)中则多达106个SPL家族成员[5,9,11-15]。
植物基因克隆是进行生命科学研究的关键组成,是进行生命科学研究时较为关键的部分[16],对于研究植物基因的表达调控模式和功能至关重要。对于已被鉴定的SPL基因家族成员,在许多植物中已被成功克隆。曾东琳等[17]以“改良香菇”芥蓝(Brassica oleracea var.alboglabra)叶片cDNA为模板,克隆得到了“改良香菇”芥蓝与叶片发育有关的SPL基因,BoSPL3-1、BoSPL10-2和BoSPL11-2;任子政等[18]以森林草莓(Fragaria vesca)的cDNA为模板,特异扩增出了森林草莓FvSPL基因;张红雨等[19]利用枸杞(Lycium barbarum L.)花药cDNA,进行克隆得到枸杞LbSPL6基因;王俊文等[20]利用“宁杞1号”枸杞为研究材料,克隆得到了枸杞花发育有关的LbSPL12基因;吴鸿飞等[21]在花芽分化期的“堰虹桂”桂花(Osmanthus fragrans)中,克隆得到了10个桂花SPL基因,并命名为OfSPL1A、OfSPL1B、OfSPL5、OfSPL6、OfSPL7、OfSPL8、OfSPL10、OfSPL11、OfSPL12、OfSPL13。
4 SPL转录因子的表达调控研究
研究表明,SPL转录因子一方面受其上游miRNA156/157调控,另一方面通过结合下游基因启动子区域的顺式作用元件,调控下游基因的表达[22]。
miRNA156/157- SPL为植物保守的基因途径,调控着植物生长发育的许多过程和性状, miR156/157 是小分子核糖核酸(microRNA),可通过介导SPL 靶基因 mRNA的切割或翻译抑制,负调控 SPL 基因功能[23]。例如对在黄瓜(Cucumis sativus L.)营养生长时期miR156/157-SPL可调控相关表型的发生,并可能在葫芦科(Cucurbitaceae)植物营养生长的早期发育中行使特殊功能[24]。在大豆[Glycine max (Linn.) Merr.]的17个含有miRNA156/157识别位点的SPL基因中,有2个能够被miRNA156通过转录剪切调控,有15个能够被miRNA156通过翻译修饰调控[25]。在番茄(Solanum lycopersicum)中,SPL基因在茎尖、花序和果实中均高表达,在幼苗、根和叶中低表达,而miR156/ 157则在幼苗、根和叶中高表达,在茎尖、花序和果实中低表达[26]。在马铃薯(Solanum tuberosum L.)Stu-miR156过表达植株中,Stu-miR156的表达量在根、茎、叶中均上调,StSPL9均下调;在其Stu-miR156STTM沉默表达植株中,Stu-miR156表达量均下调,StSPL9均上调,且二者共同调控着马铃薯的植株高度和侧根表型[27]。总而言之,miRNA156/157在SPL的表达调控中具有重要作用,且在大多数含有miRNA156/157识别位点的SPL转录因子中, miRNA156/157与SPL的表达水平在植物的生长发育过程中基本都呈负相关[8]。