彩叶树种叶色影响因素及光合特性研究进展
作者: 张阳 钱多 马喆
摘要 彩叶树种以丰富的色彩和稳定的观赏期,改善了园林景观缺乏色彩的现状,为园林景观建设提供更多选择,具有极大的发展潜力。综述了外在环境条件、叶片结构和叶绿素、花色素苷合成代谢等因素对彩叶树种叶片呈色的影响以及彩叶树种光合特性的相关研究成果,对进一步开展彩叶树品种的选育和推广栽培进行了展望,以期为彩叶树种品种改良研究、树种资源合理配置提供参考。
关键词 彩叶树种;叶片呈色;环境因子;色素代谢;光合特性
中图分类号 S 687 文献标识码 A
文章编号 0517-6611(2022)20-0012-06
doi:10.3969/j.issn.0517-6611.2022.20.004
Research Advances in Leaf Coloration Mechanism and Photosynthetic Characteristics of Colored Leaf Tree
ZHANG Yang,QIAN Duo,MA Zhe
(Beijing Vocational College of Agriculture, Beijing 102442)
Abstract Colored leaf tree is plant that has various colors and long color duration. It has great application prospects, which can solve the problem of lack of color in urban landscape and provide more high-quality choices for the construction of landscape. In this review, we summarize recent advances in the mechanism of leaf coloration and photosynthetic characteristics, including the effects of environmental factors, leaf structure, biosynthesis of chlorophyll and anthocyanin, photosynthetic efficiency and photoprotection capacity. These results will provide references for variety improvement, new variety, tree selection and planting technology.
Key words Colored leaf tree;Leaf coloration;Environmental factor;Pigment metabolism;Photosynthetic characteristics
彩叶树种是指叶片能够在生长季或生长季某一阶段稳定表达绿色以外颜色的木本植物,具备一致的叶片呈色期、落叶期及较长的观赏期[1-2]。随着我国社会经济不断发展,大众对美好生活的向往逐渐体现在精神文化层面,对高水平园林的追求愈发迫切。过去的园林景观彩叶植物品种匮乏,色彩单一缺乏层次。近年来,通过研究人员不断地引种和繁育,越来越多的彩叶树种出现在大众视野,其凭借色彩鲜艳、观赏期长、空间利用充分的特点,逐渐获得景观设计师的青睐,用于打造色彩艳丽、富于变化的园林视觉效果[3]。
彩叶树种的引种栽培具有一定的局限性。特别是我国幅员辽阔,纬度跨度广,距海远近差距较大,气候复杂多样,不同地区的环境差异及栽植条件都会影响彩叶树种的生长状况和观赏效果。该研究对彩叶树种叶色影响因素和光合特性进行总结和分析,以期为彩叶树种的选育、引种、园林应用推广等方面提供理论参考。
1 彩叶树种类型及叶片色素
彩叶树种可从叶片的颜色、色彩分布情况、呈色时期等方面进行分类。依据叶面着色方式,可分类为单色叶、双色叶、斑色叶、彩脉叶等[4]。依据叶片呈色时期则常划分为春色叶、秋色叶和常色叶。春色叶树种,如紫叶海棠、红叶石楠和香樟等春季嫩叶异色的树种;秋色叶树种,如银杏、白蜡、元宝枫等秋季叶片变色的树种;常色叶树种,如紫叶小檗、紫叶李等叶片全年保持异色状态的树种[5]。
彩叶树种的叶色由其内含色素的种类、含量及分布特征直接决定[6]。植物叶片中主要包含有三大类色素,即叶绿素、类胡萝卜素和花色素苷。叶绿素和类胡萝卜素是光合色素,是进行光合作用的重要物质。叶绿素分为叶绿素a和叶绿素b,分别呈现蓝绿色和黄绿色,类胡萝卜素分为胡萝卜素和叶黄素,呈现橙黄色和黄色[7]。一般情况下植物绿色叶片中的叶绿素占主导地位,当叶片内的类胡萝卜素相对含量升高时,会逐渐表现为黄色。金叶复叶槭黄色叶片的内外层明亮度、黄色程度与类胡萝卜素相对含量呈极显著正相关,与叶绿素含量呈极显著负相关[8]。金叶梓树、金叶接骨木、金叶榆叶片中叶绿素与类胡萝卜素含量的比值(Chl/Car)降低时,黄色程度增高,绿色程度降低[9]。转色期的黄色系榉树叶片类胡萝卜素相对含量较绿色系、红色系榉树高,为24.7%~35.4%[10]。秋季银杏叶微黄至全黄期间叶绿素总量降低了96%,类胡萝卜素只降低了66%,Chl/Car从3.403下降到0.191[11]。洒金柏黄色叶片各色素都比绿色叶片少,但类胡萝卜素与总叶绿素含量比值(Car/Chl)可达绿叶的4倍[12]。金边杂种胡颓子叶片黄化部位叶绿素比野生种低98%[13]。
花色素苷又称花青苷,是一种类黄酮类多酚化合物,由不稳定的花青素发生糖苷化形成。植物中的常见花青素依据结构不同分为6类:矢车菊素(cyanidin)、飞燕草素(delphinidin)、天竺葵素(pelargonidin)、芍药花素(peonidin)、锦葵素(malyidin)和矮牵牛素(petunidin)[14]。这6类花青素的羟基与糖类结合生成相应种类的花色素苷。花色素苷是彩叶树种叶色表达的重要物质[15],特别在红、紫色叶树种中,花色素苷的种类及相对含量是叶色变化的标准。拧筋槭[16]、榉树[17]、紫叶紫薇[18]叶片红色程度与花色素苷相对含量呈显著正相关。红花槭“艳红”叶片变红后,花色素苷含量上升为变红前的3.35倍,矢车菊素类花色素苷占总花色素苷含量的95%[19]。红叶石楠上位叶由红转绿期间,20 d内花色素苷含量下降了43.5%,叶绿素和类胡萝卜素的含量无明显波动[20]。5种北美栎树在秋冬变色期,红叶类型的纳塔栎、舒玛栎、水栎、沼生栎叶片的花青素含量均显著增加,黄叶类型的柳叶栎则类胡萝卜素含量显著增加且增幅最大[21]。欧洲刺柏“金叶疏枝”的叶片花色素苷相对含量与叶片明亮度、黄色程度呈极显著负相关,与红色程度呈极显著正相关[22]。
2 环境因子的影响
温度是促使彩叶树种叶色改变的重要环境因子,季节性变色树种依据自然温度变化呈现稳定的变色期。低温能导致光合色素的合成减少和降解,诱导花色素苷的合成,从而实现叶色表达。白蜡、火炬树、元宝枫、银杏在秋季转色期,叶片叶绿素含量随温度降低呈下降趋势,花色素苷、类胡萝卜素相对含量升高[23]。秋季日最低温低于12 ℃时,栾树叶片开始变黄,低于8 ℃时, 叶片已近全黄[24]。5 ℃低温处理欧洲刺柏“金叶疏枝”,叶片显红色相,温度上升后,叶色渐绿,明亮度、鲜艳度提高[25]。较低温度(15 ℃)处理紫叶稠李,叶片花色素苷含量相比自然环境增加3倍,花色素苷合成相关酶的活性高于自然环境条件[26]。-10 ℃低温下香樟叶肉细胞的叶绿体膜系统结构解体,部分叶绿体解体崩溃,叶绿素合成功能丧失[27]。紫叶李叶片在32 ℃高温下,彩色度降低,出现“高温返青”现象[28]。欧洲卫矛叶片花色素苷在温度≤35 ℃时比较稳定,超过35 ℃后,花色素苷溶液吸光值逐步升高,叶片颜色由鲜红向深红转变[29]。
光在植物生长发育方面起到重要作用,能改变植物色素相关关键酶的活性进而调控叶色[30]。低温无光条件下,5种彩叶杨叶片中苯丙氨酸解氨酶(PAL)、查耳酮黄烷酮异构酶(CHI)、二氢黄酮醇-4-还原酶(DFR)、类黄酮糖基转移酶(UFGT)活性均处于较低水平,花色素苷合成速率较低,通入光源后均显著上升[31]。黄金枸骨遮荫处理6 d后,叶绿素酶(CLH)和脱镁螯合酶(MCS)活性都显著下降,叶绿素积累量增加,叶片由黄转绿[32]。遮荫处理的多个品种变色期欧洲鹅耳枥,叶绿素分解速度降低,叶片延缓变黄[33]。糠椴、紫椴在遮光条件下,叶片内叶绿素总量上升,遮光程度与叶绿素a和叶绿素b的比值(Chla/Chlb)呈负相关,叶片光泽、绿色程度、黄色程度随遮光程度下降[34]。光质不同也能够影响色素含量发生变化,但相关研究有待进一步验证。美国红栌叶片花青素含量在红、黄、蓝、绿4种光质处理下均低于自然光对照组,红光和绿光处理使叶绿素含量高于对照组[35]。枫香幼苗叶片在红光处理下,叶绿素含量升高,花青素含量降低,蓝光处理则相反[36]。而范皓月[37]的研究中,则是蓝光和绿光促进红叶腺柳上部叶片叶绿素合成和积累,加速叶片红色消退向绿色转变。
水是植物细胞的主要成分,直接影响树木正常生长。中重干旱胁迫使红花檵木叶片花色素苷含量分别下降了35.18%和51.85%,叶片转绿[38]。金叶复叶槭和金叶接骨木在25%田间持水量下,叶片亮度和黄色程度降低,叶绿素总量升高197.56%和19.17%,叶绿素b增加量显著高于叶绿素a,分别上升766.67%和40.74%[39]。施加外源物质可以提供植物所需的营养成分或改变酶活性,从而调节叶片色素比例影响呈色。紫叶稠李施入氮肥45 d后,各处理叶片叶绿素含量均高于无肥料的对照组,最高处理组平均上升38.25%[40]。紫叶锦带花氮肥内加入1.1%K2SO4溶液,秋季叶片花色素苷含量比单一氮肥组增加147.17%[41]。50 mmol/L NaCl处理金叶银杏幼苗可促进叶片内类胡萝卜素的积累[42]。外源喷施0.2% FeSO4可使“砀山酥梨”黄化叶片复绿,发现类囊体部分基粒断裂片层重新连接,片层结构逐渐清晰,叶绿素总量上升[43]。pH 6柠檬酸缓冲液酸化土壤,红枫叶片内叶绿素酶(CLH)、苯丙氨酸解氨酶(PAL)活性显著上升,花色素苷含量显著增加,叶片提前转红且红色程度增强[44]。紫红叶鸡爪槭[45]、北美红枫[46]、紫叶紫薇[18]、北美枫香[47]都发现在叶片转色期可溶性糖与花色素苷呈正相关。培养基内缺少蔗糖会降低紫叶白桦组培苗花青素积累,蔗糖超过一定浓度后,花青素含量随蔗糖浓度增加而降低[48]。叶面喷施蔗糖的北美红枫叶片内可溶性糖含量和花色素苷含量高于对照组,光合色素合成减少且叶片pH降低,利于叶片呈色但影响不显著[46]。
3 叶片形态结构变化
叶绿素主要分布于叶绿体的类囊体膜中,类胡萝卜素分布于叶绿体和有色体的膜结构之中[49],叶绿体的结构变化会造成光合色素合成和积累受阻,进而影响叶片呈色。银杏变异株黄叶细胞内聚集大量液泡,叶绿体形状多不规则,叶绿体膜模糊,片层结构疏松分散、垛叠程度较低,有排列密集的嗜锇颗粒[50]。紫叶小檗叶绿体体积比小檗小,基粒垛叠疏松片层厚,嗜饿颗粒较多,同化淀粉数量多、体积大[51]。洒金柏春季顶梢黄叶叶绿体与基部绿色叶片相比发育滞后,结构不完整且基质片层排列混乱,淀粉粒少,秋季时叶绿体解体较早[52]。叶绿体主要存在于叶片栅栏组织细胞内,叶片结构也与叶绿素含量有关。枫香变色末期红色逐渐加深,叶片栅栏组织、海绵组织以及栅海比逐渐减小,叶绿体扁长形,嗜锇颗粒逐渐增大[53]。金叶金钟花叶片较薄,栅海比极显著小于原种朝鲜金钟花,并且通过观察超微结构,发现其叶绿体呈球状或锥状,被膜模糊且破损严重,发育基本停滞在质体阶段,类囊体无基粒和基粒片层[54]。
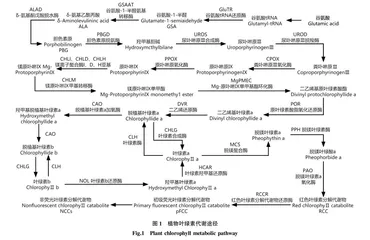
4 相关基因表达调控
色素的合成和表达是多步骤、多酶参与的复杂生物学过程,改变相关基因的表达会造成色素代谢紊乱。结构基因直接编码色素代谢过程相关酶[55],彩叶香樟叶片中的叶绿素积累缓慢,其谷氨酰t-RNA还原酶基因(CcHEMA)、谷氨酸酯-1-半醛-2,1-氨基变位酶基因(CcGSA)、胆色素原脱氨酶基因(CcHEMC)、尿卟啉原Ⅲ合成酶基因(CcHEMD)、粪卟啉原Ⅲ氧化酶基因(CcHEMF)、镁离子螯合酶H亚基(CcCHLH)、I亚基(CcCHLI)、D亚基(CcCHLD)的表达量皆低于野生香樟[56]。中红杨、全红杨[57]、金心黄杨、金边黄杨[58]对比绿叶品种,均发现粪卟啉原Ⅲ(CoprogenⅢ)形成原卟啉Ⅸ(ProtoⅨ)的过程受阻。淡黄叶水稻叶绿素合成过程在原卟啉Ⅸ(ProtoⅨ)形成镁原卟啉Ⅸ(Mg-ProtoⅨ)步骤受阻,正常野生绿叶的镁离子螯合酶活性是淡黄叶的5.65倍[59]。舒马栎黄色叶片中Qs CHLD基因的表达水平仅为绿叶品种的60%[60],与沉默豌豆[61]CHLD获得黄叶植株的结果相一致。银杏金黄色叶突变体中叶绿素降解相关的基因NYC、NOL及类胡萝卜素生物合成相关15-顺式-ζ-胡萝卜素异构酶(Z-ISO)、ζ-胡萝卜素脱氢酶(ZDS)、番茄红素ε-环化酶(LCYE)均上调表达[62]。植物叶绿素代谢途径见图1。