入侵杂草薇甘菊叶不同发育期微形态特征研究
作者: 曹畅 黄梦婷 何志敏 罗肖莹 吴江兰 王长宝
摘要[目的]揭示薇甘菊叶不同发育时期微形态学特征,为深入了解该入侵物种的生理特性及生态适应性提供重要参考。[方法] 利用临时装片法,在光学显微镜下观测叶表皮细胞、气孔器及表皮毛的形态性状。[结果]叶长2.90~6.05 cm时,上表皮细胞面积和长宽随着叶生长呈现稳步上升趋势,叶长6.50~14.00 cm时,上表皮细胞面积和长宽变化相对较小,总体上表现为缓速增长;下表皮细胞随叶生长变化较小,但长宽比均值与上表皮差距很小;气孔器主要发生在下表皮,叶长对气孔器面积比的影响较小,气孔器的长宽及长宽比没有随发育期的变化出现明显波动;展叶期上表皮毛个数大于下表皮,随着叶的发育成熟,迅速减少。[结论]薇甘菊叶的不同发育期表皮微形态特征受环境因素的影响,反映叶片发育与其功能相适应。
关键词薇甘菊;表皮细胞;气孔器;表皮毛
中图分类号S451文献标识码A
文章编号0517-6611(2022)13-0102-04
doi:10.3969/j.issn.0517-6611.2022.13.027
Study on Micromorphological Characteristics at Different Developmental Stages of Invasive Weeds Mikania micrantha Leaf
CAO Chang, HUANG Meng-ting, HE Zhi-min et al(College of Biology and Pharmacy, Yulin Normal University, Yulin, Guangxi 537000)
Abstract[Objective]To reveal the micromorphological characteristics at different developmental stage of Mikania micrantha leaf, which provides an important reference for further understanding of the physiological characteristics and ecological adaptation of this invasive species. [Method] The morphological features of epidermal cells, stomata and epidermal hair were observed under light microscopy using temporary slide method. [Result]The area and length of adaxial epidermal cell increased steadily during leaf length 2.90 cm to 6.05 cm, while between 6.50 cm to 14.00 cm, the epidermal cell area changed relatively little. The abaxial epidermal cells change less with leaf growth, and the difference of the length/width between the adaxial and abaxial epidermis was very small. The stomata mainly occur in the adaxial epidermis, and the leaf length has less effect on the area ratio of stomata. The length, width and length/width of stomata does not fluctuate significantly in different development period. The number of epidermal hairs in the adaxial epidermis is greater than that of the adaxial epidermis, and decreases rapidly as the leaves mature. [Conclusion]Environmental factors influence the leaf micromorphological characteristics of Mikania micrantha, which reflects that the leaf development is adapted to its function.
Key wordsMikania micrantha;Epidermal cell;Stomatal;Trichome
薇甘菊(Mikania micrantha H.B.K)原产于南美洲和中美洲,现已广泛传播到亚洲热带国家,成为当今世界热带、亚热带地区危害最严重的杂草之一[1-2]。1919年薇甘菊作为杂草在中国香港出现,1984年在深圳发现[3-5]。广西最早证实薇甘菊是2008 年在陆川县铁路沿线发现,目前,桂东南玉林市管辖的陆川县、博白县、北流市和容县均有记录[6]。笔者在玉林市区部分公园和校园绿化带也发现有薇甘菊的分布,说明该种正在适应城市人工干扰环境。植物进化过程中,叶片形态对环境变化较为敏感,能体现环境因子的影响以及对环境的适应性[7]。薇甘菊的抗强光胁迫能力并不强,但是它很快长出新叶,能够很好地利用强光资源进行光合作用,同时还可以通过调节自身叶片的形态特征来维持其对不同光照环境的入侵能力[8],表明其叶片具有很强的形态和功能可塑性。已有研究发现,薇甘菊幼苗初期生长缓慢,在30 d内苗高仅11 cm,单株叶面积0.33 cm2,但随着苗龄的增长,其生长随之加快,茎部产生不定根,并迅速扩展。这种快速生长的结构基础来源于叶柄、叶脉、茎等非同化器官,具有C4植物的典型维管束鞘结构特征以及气孔的调节功能[9-10]。虽然,前人开展了薇甘菊成熟植物体茎叶解剖特征研究,但对其叶的微形态特征认识尚不清楚。笔者通过对薇甘菊叶不同发育期的表皮细胞、气孔器和表皮毛的特征进行了比较分析,为进一步了解薇甘菊叶片的环境适应性提供参考资料。
1材料与方法
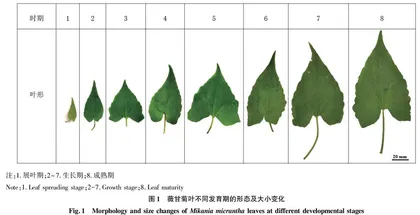
1.1试材采集选取玉林师范学院东校区挂榜山马尾松林下小路边薇甘菊,进行叶片生长情况观察,每3 d观察1次,测量叶片大小并拍照记录数据。经过60 d的观测,掌握薇甘菊形态大小的时间变化规律,由此将薇甘菊叶从展叶期到成熟期按照大小划分为8个阶段进行取样(图1)。试验材料的采集时间均为晴天14:00—16:00。
1.2试验方法
1.2.1叶表皮细胞观测。每组采取2片新鲜叶子,双面刀片刮除叶肉,清水冲洗后经醋酸洋红染色,制成临时装片镜检,选择5个清晰的视野在Olympus显微镜下观察,利用CELL软件绘制细胞轮廓并进行数据测量,每个视野测20个细胞的长宽(长宽比=细胞长/细胞宽)及面积,每组共200个细胞。观察结束后拍照保存,并分析表皮细胞所属的形态类型。
1.2.2气孔器观测。每组选择40个气孔器测量长、宽和面积后取平均值,再选择6个视野单位面积,计算气孔器面积、气孔器长、宽以及长宽比值、气孔器面积与视野单位面积比值。气孔器面积与视野面积比值=视野内气孔器面积/视野面积;气孔器长宽比值=保卫细胞长/保卫细胞宽。
1.2.3表皮毛观测。每组分别测量视野内上表皮单位面积下的表皮毛个数(单位面积为820 800 μm2),上下表皮各选取5个不同位置的视野,统计表皮毛的个数,取平均值,并计算每1万μm2的表皮毛个数。每1万μm2的表皮毛个数=(视野内表皮毛个数/视野面积)×104。
2结果与分析
2.1薇甘菊叶不同发育期表皮细胞特征由表1可知,叶长2.90~6.05 cm时,上表皮细胞面积和长、宽均值呈上升趋势,但个体细胞生长不同;叶长6.50~14.00 cm时,上表皮细胞面积和长、宽均值变化相对较小,总体上表现为缓速增长。上表皮细胞各生长阶段的细胞面积和长、宽均值大多大于下表皮,对于下表皮细胞来说,薇甘菊叶长2.90~14.00 cm,表皮细胞面积并没有因为叶的生长而出现大幅变化,总体趋近于200~300 μm2,长、宽均值也较上表皮略小,但长宽比均值上下表皮都很接近。
薇甘菊上表皮细胞垂周壁样式多呈直状,少数呈波状,细胞形态多呈多边形和无规则形,且垂周壁波状和细胞无规则形、垂周壁平直状和细胞多边形同时出现(表1、图2A、C)。下表皮细胞垂周壁样式全部呈波状,细胞形态全部呈无规则形,且细胞垂周壁和细胞形态的变化较上表皮小(表1、图2B、D)。
2.2薇甘菊叶不同发育期气孔器特征薇甘菊上下表皮均有气孔器,但上表皮气孔器偶有出现,故气孔器的观测均采用下表皮进行。薇甘菊的气孔器由2个肾状保卫细胞围合而成,中间留有大小不同的气孔,均匀分散在叶面上,镶嵌在无规则的表皮细胞间(图2B、D)。由表2可知,叶长对气孔器面积比影响较小,气孔器的长宽以及长宽比也没有随着发育期的变化出现明显的波动,长宽比均处于一个平均值范围内(1.39~1.66),面积较大的气孔器长宽略大于面积较小的气孔器(表2)。
2.3薇甘菊叶不同发育期表皮毛特征薇甘菊叶上下表皮均有表皮毛,其中展叶期上表皮毛密度大于下表皮(图2E、F、表3),此时表皮毛相较于成熟期密集。当薇甘菊叶进入生长期,随着叶片面积的增加和成熟度的上升,开始不断脱落表皮毛尤其是上表皮到了成熟叶后期,表皮毛的个数极小,仅0.032个,说明叶肉细胞和叶表皮已经发育完全,不再依赖表皮毛的保护,而此时的表皮毛也相对较大,更加粗壮(图2G、H)。
3讨论与结论
作为一种入侵作物,薇甘菊能够迅速生长发育与其叶片的结构迅速成熟密切相关。薇甘菊幼叶期下表皮的气孔器和表皮细胞已经趋向于成熟,与成熟期叶片细胞面积以及细胞长宽并无较大区别,这些成熟的细胞促进了薇甘菊的迅速生长,这与张衷华[9]的结论相一致。幼叶细胞结构在30 d内的快速构建为薇甘菊生长发育期第1次快速增长打下了坚实的基础。展叶期的叶片尚未发育完全,强烈的太阳光会灼伤叶组织,薇甘菊通过形成大量表皮毛反射太阳光而保护靠近上表皮的叶肉组织,从而保证其顺利度过幼叶期,有利于在叶面上下表皮之间形成栅栏组织和海绵组织,促进上表皮扁平的细胞迅速生长,从而能够适应太阳光强的程度[11]。展叶期结束后,随着叶片的生长,叶片开始脱落大量表皮毛,从而增强了气孔器的气体交换和表面光照的吸收,更有利于植物本身的营养补给,当叶片正式进入成熟期时,表皮毛的密度下降至最低。气孔是植物吸收CO2、散失水分的主要通道,是植物与外界信息交流的主要窗口,直接影响着植物的蒸腾作用、光合作用等主要生理活动,在植物的生长发育过程中起着重要作用[12]。薇甘菊叶的不同发育期下表皮气孔器面积和下表皮细胞面积数据变化趋近,这反映植物的发育是与其功能相适应的。
该试验中叶长6.50~14.00 cm时,薇甘菊上表皮细胞面积和长、宽出现少许波动变化,认为是由于光强和环境因素导致的植物个体的微小差异。从数据分析得出,部分薇甘菊叶片在发育期由于光照较少而生长缓慢,个别叶片因光照较好而生长迅速,从而影响了表皮细胞的生长,这是薇甘菊对环境变化的反映,进而验证了薇甘菊对光强有很强的环境适应性,这也与魏巍[13]的研究结果一致。该研究揭示了薇甘菊叶片不同发育时期的微形态学特征,为深入了解该入侵物种的生理特性及生态适应性提供参考。
参考文献
[1] COCK M J W.Potential biological control agents for Mikania micrantha from the Neotropical region [J].Tropical pest management,1982,28(3):242-254.
[2] ZHANG L Y,YE W H,CAO H L,et al.Mikania micrantha H.B.K.in China-An overview[J].Weed research,2004,44(1):42-49.
[3] 杜凡,杨宇明,李俊清,等.云南假泽兰属植物及薇甘菊的危害[J].云南植物研究,2006,28(5):505-508.
[4] 昝启杰,王勇军,王伯荪,等.外来杂草薇甘菊的分布及危害[J].生态学杂志,2000,19(6):58-61,77.
[5] 练惠通,黄泽文,张波,等.广东农业外来有害生物薇甘菊入侵的历史阶段与特点[J].安徽农业科学,2014,42(20):6695-6698,6755.
[6] 韦春强,潘玉梅,唐赛春,等.入侵植物薇甘菊在广西的分布与危害[J].广西植物,2014,34(6):816-820.
[7] HENRY H A L,AARSSEN L W.On the relationship between shade tolerance and shade avoidance strategies in woodland plants [J].Oikos,1997,80(3):575-582.
[8] 黄乔乔,李晓霞,程汉亭,等.不同光照及采食条件对薇甘菊和鸡屎藤叶片形态可塑性的影响[J].热带农业科学,2013,33(7):11-16.
[9] 张衷华.林业有害植物薇甘菊入侵特性的研究[D].哈尔滨:东北林业大学,2006.
[10] 王文杰,张衷华,祖元刚,等.薇甘菊(Mikania micrantha)非同化器官光合特征及其生态学意义[J].生态学报,2009,29(1):28-36.
[11] 张水珠.薇甘菊不同居群蛋白质和解剖结构比较研究[D].广州:华南农业大学,2007.
[12] 陈华,胡晓颖,沈浩,等.田野菟丝子寄生对薇甘菊气孔行为的影响[J].广西植物,2008,28(5):655-660.
[13] 魏巍,侯玉平,彭少麟,等.不同光照强度对入侵植物薇甘菊(Mikania micrantha)和飞机草(Chromolaena odorata)生长及生物量分配的影响[J].生态学报,2017,37(18):6021-6028.